 Cellemembraner: Dynamisk, Kommuniserende, Designet
Cellemembraner: Dynamisk, Kommuniserende, Designet
Evolution news; 21. mai 2018
Bilde 1. Kilde: Illustra Media, Origin: Design, Chance, og det første livet på jorden.
Oversatt herfra.
Enkle bilag av lipider -her? Nei! Cellemembraner har vist seg som mester spillere i tallrike cellulære funksjoner. Disse 'skinnene' av celler, som ikke bare omgir hele celler, men også deres kjerne og organeller, er virkelig fantastiske. Hvis du ser på ensartede 'skinn' som gummiballonger rundt celler, har du oppfattet det galt. Cellemembraner omfatter ikke bare lipidene, men kanalene, maskiner og prosesser som er innebygd i dem, akkurat som menneskelig hud er et komplekst organ som omfatter mange porer, sensorer og støttekar. Current Biology -her, viet spesielt oppmerksomhet til cellemembraner. La oss se på noen høydepunkter i de siste oppdagelsene.
Først av alt har forskere funnet ut at forskjellige lipidmolekyler kan produsere forskjellige kurver og former. Vi vet at mikrober kan anta en rekke former, fra kuler til stenger til formløse lik amøber. Figur 1 i en artikkel av Garcia et al. -her viser at "lipider kan bestemme membranskurvaturen", som i sylindrisk, konisk og omvendt konisk. Denne fleksibiliteten er viktig ikke bare for cellens overordnede morfologi, men for hver av dens innebygde strukturer. Membranene må være tøffe nok til å holde ut uønsket materiale og gi fast støtte til sitt maskineri. Sikkert nok ville ikke den raskt spinnende bakterielle-flagellum fungere i en skrøpelig konstruert membran! Og likevel trenger membraner fleksibilitet for å bøye seg og gi inngang for ønskelige molekyler. Membraner er de inngjerdede veggene der aktiv transport foregår, som overvinner den naturlige loven til osmose som ville forsumpe en celle med uønskede molekyler eller lekke ut dens kronjuveler.
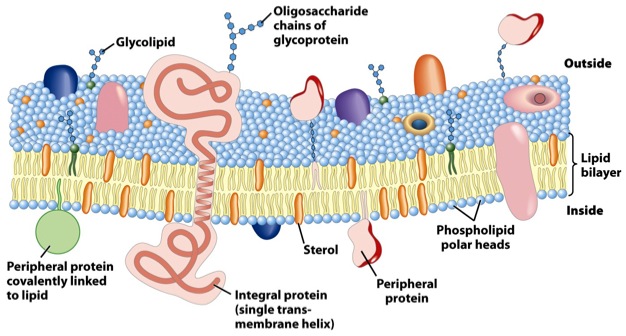 Den ciliære membranen
Den ciliære membranen
I 'Hvordan ciliær membran organiseres fra innsiden-ut for å kommunisere fra utsiden-inn,' beskriver forfattere membranene som omgir cilium (en av de irreduserbart-komplekse molekylære maskiner Michael Behe diskuterte i sin bok, Darwins Black Box tilbake i 1996). Disse viktige cellulære systemene, som ikke alle er bevegelige, fungerer som antenner i de fleste celler. Kunstige antenner er vanligvis laget av metall, og reagerer på elektromagnetiske bølger. Hva reagerer cellulære antenner på?
--Som et viktig grensesnitt med resten av verden er cilia-membranen et fascinerende eksempel på hvordan membran-spesialisering tildeler kritiske funksjoner, slik at cilium kan fungere som antenne for cellen. Nødvendig for signaleringsfunksjonene til cilia er presis kontroll av cilia-membran kategorisering, sammensetning og morfologi. Ulike typer cilia utviser mangfoldige membran-morfologier, og morfologien og remodelleringen av ciliære membraner kan dynamisk modifiseres ved signalaktivitet. [Kursivering lagt til.]
Cilia er mye mer dynamisk enn de passive metallantennene vi er vant til på menneskelig aktivitet. Behe fortalte om 9 + 2 mikrotubule innmat, med sine molekylære motorer, i sin bok, men selv membranen rundt alt det maskineriet er høyt spesialisert. Hvis du vil ha en smak av hvor sofistikert membranen rundt et cilium er, se på noen av tingene de må gjøre:
--Cilia, organeller som beveger seg for å utføre funksjoner som befruktning og signal for å utføre funksjoner som foto-mottak og embryonisk mønstergivning, består av en kjerne av ni-folds dublett mikrotubuler overkledd av en membran. Distinkte typer av cilia viser forskjellige membran-morfologier, alt fra enkle buede beholdere til de høyt utsmykkede invaginasjoner og membranskiver for ytre segmenter av fotoreceptor. Kritisk for ciliaens evne til å signalere, er både proteinet og lipid-sammensetningene av ciliære membraner forskjellige fra andre cellemembraner. Denne spesialiseringen gir en unik utfordring for cellen fordi, i motsetning til membranbundne organeller, er cilia-membranen tilstøtende med den omgivende plasma-membranen. Denne distinkte cilia-membranen genereres i samspill med flere membran-omformingshendelser som omfatter prosessen med cilio-genese. Når cilia er dannet, er kontrollen av ciliær membran-blanding avhengig av diskrete molekylære maskiner, inklusiv en barriere for membranproteiner som kommer inn i cilia i en spesialisert region i basen av cilium som kalles overgangssonen og en trafikk-tilpasser som styrer G-ProteinKoblet Reseptor (GPKR) lokalisering til cilium, kalt BBSome. Cilia-membranen kan videre omformes ved fjerning av membranproteiner ved frigjøring av ciliære ekstra-cellulære vesikler som kan fungere i intercellulær kommunikasjon, fjerning av unødvendige proteiner eller cilia-demontering.
Spirende Membranes: Det Cellulære Internett
Visste du at celler bruker e-post? Se på et fascinerende papir i serien om “Exosomes and Ectosomes in Intercellular Communication" -her. I denne artikkelen vurderer Jacopo Meldolesi hva som er kjent om 'spirende' vitenskap om intercellulær kommunikasjon.
--Inntil for nesten 30 år siden ble membranfragmenter observert i ekstracellulær væske antatt å skyldes apoptose og andre prosesser av celledød. Dette forslaget har blitt stadig større fokusert i løpet av de siste to tiårene, og er nå kjent å være feil. Faktisk inneholder ekstracellulær væske sammen med membranfragmenter to typer ekstracellulær vesikkel (EV), eksosomer og ektosomer, som virker nær og også i betydelig avstand fra deres foreldre-celler.
Denne historien er sammenlignbar med "søppel-DNA" myten. Membranfragmentene Meldolesi snakker om er faktisk pakker med informasjon som reiser mellom celler. Eksosomer er mindre (50-150 nm) og er dannet fra endocytiske membraner; ektosomer er større (100-500 nm) og dannes fra plasmamembran. Fyllt med proteiner, RNAer, enzymer og andre informasjonsmolekyler, og stuet med flere reseptorer, kobler disse reisepakkene (illustrert på Figur 2) celler til å danne et slags sosialt nettverk. Analogen med e-post er slående:
--For både eksosomer og ektosomer er overflaten og hulrom-lastene heterogene når man sammenligner vesikler som frigis av forskjellige celletyper eller av enkeltceller i forskjellige funksjonelle tilstander. Ved frigivelse navigerer de to typer vesikel gjennom ekstracellulær væske for varierende tider 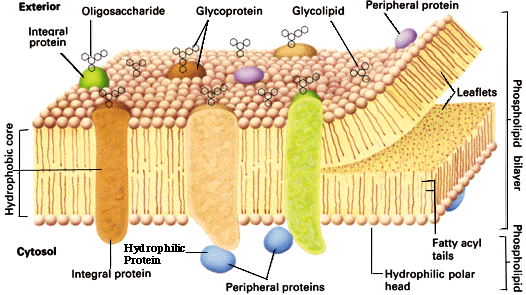 og avstander. Deretter interagerer de med anerkjente målceller og gjennomgår fusjon med endocytiske eller plasma-membraner, etterfulgt av integrasjon av vesikkel-membraner inn i deres fusjons-membraner og utslipp av hulroms-laster inn i cytosolen, noe som resulterer i endringer i cellulær fysiologi. Etter sammensmelting kan eksosom/ektosom-komponenter bli samlet i nye vesikler som deretter resirkuleres til andre celler, som aktiverer utøvende nettverk.
og avstander. Deretter interagerer de med anerkjente målceller og gjennomgår fusjon med endocytiske eller plasma-membraner, etterfulgt av integrasjon av vesikkel-membraner inn i deres fusjons-membraner og utslipp av hulroms-laster inn i cytosolen, noe som resulterer i endringer i cellulær fysiologi. Etter sammensmelting kan eksosom/ektosom-komponenter bli samlet i nye vesikler som deretter resirkuleres til andre celler, som aktiverer utøvende nettverk.
Meldolesi sier at "alle celler er nå kjent for å kommunisere ved utveksling av store molekyler via EV-trafikk", men mye gjenstår ukjent om hva disse små pakkene med informasjon faktisk gjør. Forskere har kjent om eksocytose og endocytose, prosesser som leverer molekyler fra celle til celle, som vesikler som leverer nevro-transmittere på tvers av synapser mellom nerveceller. Eksosomer og ektosomer, derimot, kan reise lange avstander og levere ulike typer molekyler til andre celler. "Ikke-kodende RNAer og DNA-sekvenser har blitt funnet blant innhooldet i eksosom hulrom," sier han, "selv om deres akkumulasjons-mekanismer ikke er klarlagt."
Til forskjell fra e-post ser disse membranbundne pakkene ut til å dele materielle fungerende deler fra celle til celle, som om du kan sende en venn noen av dine enzymer eller gener som e-post. Det fremkaller nesten tvil om celleteori i seg selv: Hvor blir det av ideen om en autonom, uavhengig levedyktig celle når naboene deler alle deler? Tallrike forskningsspørsmål stammer fra realiseringen at cellene deler informasjon på denne måten: Hvilken prosess bestemmer innholdet? Hvordan navigerer pakkene til målet? Hvilke sykdommer resulterer når ting går galt? Fremtiden virker lys for å oppdage funksjoner i disse informasjonsfylte hylster av membraner som ble antatt å være rester fra døde celler.
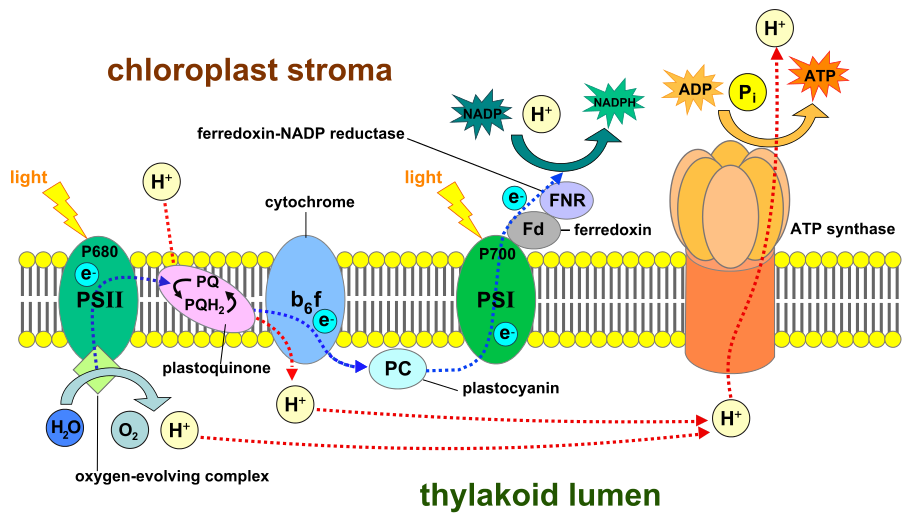 Kjernemembraner
Kjernemembraner
La oss se kort på et annet eksempel: kjernemembranen. DeMagistris og Antonin -her; beskriver "den dynamiske naturen til kjerne-hylsteret" i deres artikkel. Forbered deg på å bli forbauset:
Som andre membraner i cellen består kjernemembranen primært av lipid-bilag. Denne membranen står imidlertid overfor en stor kvalitets-kontrollutfordring. Dens doble membran må bli revet ned og ombygd til to identiske kopier ved hver celle-deling. Dette må skje raskt, uten lekkasje av sensitivt kjernemateriale i cytoplasmaen, etter at andre maskiner har jobbet febrilsk for å duplisere hele DNA, spole og supercoile det inn i kromosomer og ordnet kromosomene på en mitotisk spindel. Ikke bare det, nukleær-membranen er stufull med hundrevis av kopier av Nuklerært Pore Kompleks (NPK), én av de mest utførlige proteinstrukturer i cellen. Hver kopi av NPK har over 500 proteiner. Snakk om en massiv logistikkoperasjon!
--Eukaryoter organiserer karakteristisk sitt genom i et eget kammer, kjernen, som er omgitt av kjerne-hylster som en barriere. Brudd på kjerne-hylsteret og eksponering av kromatin truer celle-levedyktighet og forårsaker genom-ustabilitet. Til tross for den grunnleggende grensefunksjonen, gjennomgår kjerne-hylstre bemerkelsesverdige morfologiske forandringer, mest merkbare under mitose. Her oppsummerer vi vår nåværende forståelse av kjerne-hylster dynamikk og dets omskiftelige forhold til endoplasmatisk retikulum. Vi diskuterer hvordan kjerne-hylsteret omformes for å sette inn kjernepore-komplekser, transportportene til kjernen, inn i sin doble membranstruktur. Nylige 3D-elektronmikroskopi-tidskurs for montering av kjerneporekomplekser viser at disse strukturene integreres i kjerne-hylsteret under interfase og mitose ved å følge ulike veier. Begge banene sikrer at porene dannes i det kjerne-hylsteret som forbinder cytoplasma og nukleoplasma.
Figur 1 i artikkelen viser hvordan kjerne-hylsteret er tilstøtende med endoplasmatisk retikulum (ER). Figur 2 viser hvordan ER posisjonerer NPKene for innføring i membranen og monterer dem i trinn. Detaljer om stadiene er vist i figur 3, med de avsluttende trinnene av samlingen vist i figur 4. Husk at disse er forenklede diagrammer. Kunne vi se den faktiske operasjonen i den skalaen, ville vi bli blåst overende av dens effektivitet og hurtighet. En ting vet vi; celledeling fungerer vanligvis uten om og men.
 Konklusjoner
Konklusjoner
Alle artiklene i serien er fascinerende: du kan lære om caveolae, eisosomer, fusogener, multisubunit-teter og andre strukturer i cellemembraner. Det er som å lære om svettekjertler, temperatursensorer, berøringssensorer, hårsekk og alle underverkene som er innebygd i menneskelig hud for første gang etter å ha tenkt på hud som et formløst, ensartet vev. Og du kan lære om membraner og evolusjon. Evolusjon?
I en av artiklene håndterer Sven B. Gould " Membranes and Evolution." -her. Dette burde være bra. Når alt kommer til alt har vi sett frem til dette punktet, hvordan skal darwinismen håndtere dette? Gould vet hva han står overfor:
--Biologiske membraner er tynne amfifile kapper, bare noen få nanometer tykke, som både definerer grensen til alle cellene, samt mangfoldet av de indre delene i eukaryoter. Plasma-membranen til en typisk prokaryote inneholder ca. 20-30% av cellens uttrykte proteiner, og lipidene utgjør ca. 10% av cellens tørre masse. Tallene for eukaryote celler er sammenlignbare - differansen i overflateareal til volumforholdet kompenseres totalt av det eukaryote endomembran-systemet.  Omtrent en fjerdedel av proteinet kodet av det humane genomet bærer i det minste et strekk av en sekvens som antas å tjene som et transmembran-domene. Membraner er verter for substratutveksling, sensing og kommunikasjon, og livgivende energibesparelse via kjemi-osmotisk ATP-syntese.
Omtrent en fjerdedel av proteinet kodet av det humane genomet bærer i det minste et strekk av en sekvens som antas å tjene som et transmembran-domene. Membraner er verter for substratutveksling, sensing og kommunikasjon, og livgivende energibesparelse via kjemi-osmotisk ATP-syntese.
Han er ikke ferdig; Han noterer seg også at "En eukaryotisk celle kan huse flere hundre forskjellige typer lipidarter." Hvordan i all verden skal han håndtere denne utfordringen ved å bruke bare blinde, ikke-styrte krefter?
I hovedsak er svaret hans: "Det utviklet seg." Seriøst; han hevder bare at evolusjonen gjorde alt:
--Dette mangfoldet er et produkt av omtrent fire milliarder år med evolusjon; som begynner med opprinnelsen til prokaryotisk liv, gjennom splittelsen i bakterier og arkea, opprinnelsen til eukaryoter for to milliard år siden, og frem til i dag. Ved eukaryo-genese (overgangen fra prokaryot til eukaryotisk liv) økte cellekompleksiteten med fremveksten av nye membranrom, hvor mitokondrion tok senterstadiet. Membrankompleksiteten nådde en topp gjennom sekundære - og etterfølgende tertiære endo-symbioser, som skaper organeller av eukaryotisk opprinnelse omgitt av mer enn to membraner. Det evolusjonære 'klatre fra enkle til komplekse' celler er først og fremst assosiert med en økning i membranbundne og spesialiserte reaksjonskamre. Siden flukten fra hydrotermiske ventiler separerte en kontinuerlig arvelighet av plasmamembraner livet fra døden og en plasmatisk fra en ikke-plasmatisk fase.
Hvordan starter en med å svare på det? En bruker 'opprinnelsen til' og 'fremveksten av' lik magiske ord, selv for molekylære motorer som ATP-syntase. Kanskje det beste svaret ville være å bruke lignende ordlyd med noe vi kjenner til: 'evolusjonen' av biler eller arkitektur eller fabrikker. Men ta først ut de designende sinnene. Se så om forklaringen er i stand til å passere gjennom sinnets membran.
Oversettelse og bilder ved Asbjørn E. Lund